Czasopisma
Wszystkie czasopisma
Advances in Dermatology and Allergology/Postępy Dermatologii i Alergologii
Advances in Interventional Cardiology/Postępy w Kardiologii Interwencyjnej
Anaesthesiology Intensive Therapy
Archives of Medical Science
Biology of Sport
Central European Journal of Immunology
Folia Neuropathologica
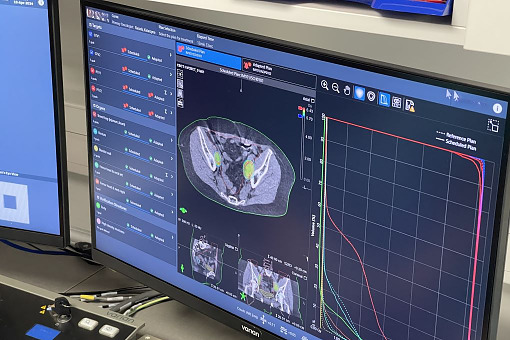
Journal of Contemporary Brachytherapy
Pediatric Endocrinology Diabetes and Metabolism
Pielęgniarstwo Chirurgiczne i Angiologiczne/Surgical and Vascular Nursing
Polish Journal of Pathology
Prenatal Cardiology
Videosurgery and Other Miniinvasive Techniques






















 Rola rekombinowanych czynników wzrostu w prewencji gorączki neutropenicznej
Rola rekombinowanych czynników wzrostu w prewencji gorączki neutropenicznej
 eOnkologiaNews
eOnkologiaNews
 Współczesna Onkologia
Współczesna Onkologia
 Journal of Contemporary Brachytherapy
Journal of Contemporary Brachytherapy






